Evolution never sTOPS!


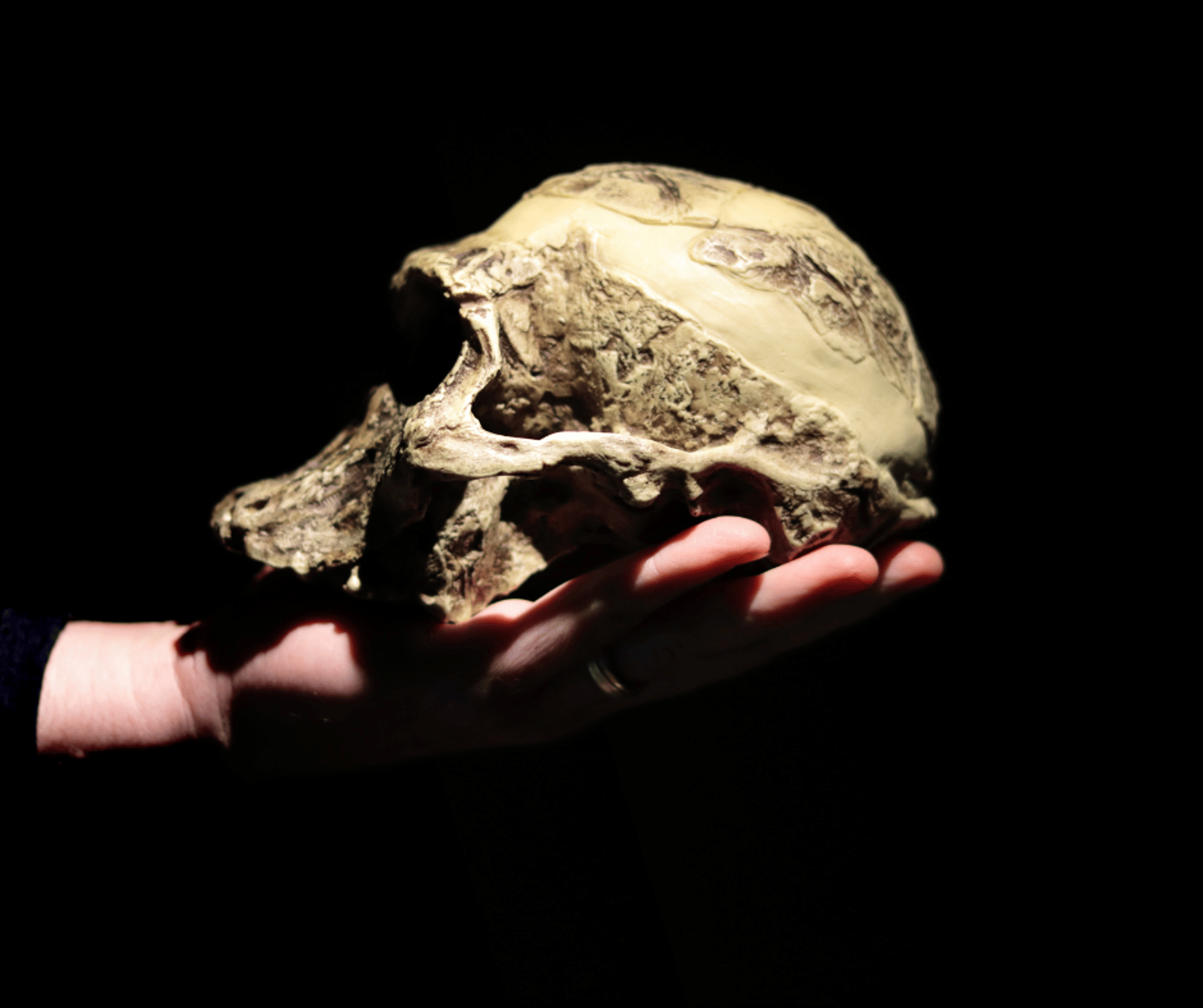
"Revisione" paleobiologica del genere 𝘼𝙪𝙨𝙩𝙧𝙖𝙡𝙤𝙥𝙞𝙩𝙝𝙚𝙘𝙪𝙨
In questi anni sono state rinvenute molte specie associate ed attribuite al genere 𝘼𝙪𝙨𝙩𝙧𝙖𝙡𝙤𝙥𝙞𝙩𝙝𝙚𝙘𝙪𝙨, ma molti di queste potrebbero non essere valide.
IL GENERE 𝘼𝙪𝙨𝙩𝙧𝙖𝙡𝙤𝙥𝙞𝙩𝙝𝙚𝙘𝙪𝙨
9/4/202513 min leggere
In questi anni sono state rinvenute molte specie associate al genere Australopithecus, da cui si origineranno i generi Homo e Paranthropus. I ricercatori dello studio cercano in qualche modo di fare una sorta di "revisione" paleobiologica su questo gruppo in quanto è importantissimo per capire come il genere Homo si sia originato, e soprattutto perché le nostre conoscenze sono ormai cambiate. Per esempio, le due principali specie (Australopithecus afarensis ed Australopithecus africanus) erano precedentemente descritte come creature bipedi incapaci di utilizzare strumenti in pietra quando, in realtà, erano anche arboricoli (potevano scegliere la tipologia di locomozione in base alle proprie esigenze). La loro dieta era variegata ed includeva prodotti alimentari a base vegetale C3 e C4, integrati da risorse animali attraverso strumenti litici (Lomekwi, 3,3 milioni di anni circa). Bene, dopo questo preambolo, possiamo analizzare punto per punto i risultati della ricerca.
Morfologie e diversità delle specie
Si esclude in primis il genere Paranthropus in quanto si tratta di un genere monofiletico, pertanto le specie da considerare in questo discorso sono ben 8: A. afarensis, A. africanus, A. garhi, A. sediba, A. deyiremeda, A. bahrelghazali, A. anamensis e A. prometheus (quest'ultima specie già da molti anni non viene considerata come tale, ma gli autori della ricerca la prendono comunque in considerazione). Le prime 4 specie, senza entrare troppo nei dettagli, possiedono morfologie condivise e derivate (uniche per ogni specie) e ciò permette di considerarle valide, mentre le altre quattro non presentano prove solide.
A. africanus presenta uno scheletro quasi completo e, come indicano molti dati, potrebbe essere l'antenata di A. sediba, anche se si differenzia per molti tratti cranici e post-cranici; A. garhi, invece, sembrerebbe essere diversa da A. afarensis, la sua potenziale antenata, in quanto presenta caratteristiche simili al genere Paranthropus, come ad esempio una debole cresta trasversale sul terzo premolare della mascella e un canino caratterizzato da una grande corona. Vediamo tutte le situazioni:
A. bahrelghazali potrebbe essere considerata come appartenente ad A. afarensis;
A. deyiremeda potrebbe essere una specie separata, anche se abbiamo bisogno di più prove. È una specie simile ad A. afarensis per via della dimensione mandibolare, ma differisce da essa per la posizione del processo zigomatico, per l'assenza di seni venosi occipito-marginali e per via di un piccolo meato acustico esterno e di una faccia piatta. Il risultato è che il materiale di A. deyiremeda potrebbe appartenere ad una popolazione di A. afarensis e, come tale, presenterebbe una certa variabilità intraspecifica;
Esiste anche il "problema" Kenyanthropus platyops, una specie considerata tale solo attraverso pochi e frammentati reperti cranici e che pare non somigliare ad A. afarensis. Le differenze morfologiche potrebbero essere il risultato di deformazioni tafonomiche, ma le successive scoperte indicano che queste due specie non appartengono alla stessa popolazione per via del diverso adattamento dietetico di A. afarensis. Tuttavia, lo studio dei reperti mascellari e mandibolari di BRT-VP-3/1 (olotipo di A. deyiremeda) ha ridotto il divario tra A. afarensis, K. platyops e A. deyiremeda, pertanto in futuro potremmo assistere alla comparsa di Australopithecus platyops;
Anche A. anamensis parrebbe essere una specie separata da A. afarensis, ma le somiglianze sia con gli individui giovanili che con gli "anziani" di Laetoli di A. afarensis sembrano affossare quest'ipotesi. Il cranio di A. anamensis, denominato MRD-VP-1/1 e antico 3,8 milioni di anni circa, presenta sia caratteri plesiomorfi (antichi) che derivati, ma questi cambiamenti graduali e a "mosaico" si osservano maggiormente sulla mandibola e sui denti. I tratti plesiomorfi includono un canino superiore relativamente grande e allungato mediodistalmente, allineato con la fila dei denti post-canini, un piccolo meato acustico esterno, un margine nasale laterale arrotondato e un palato relativamente ristretto, mentre il postcranio somiglia a quello di A. afarensis. Quindi, gli elementi cranici, mandibolari e dentali e le condizioni "primitive" di A. anamensis e quelle derivate di A. afarensis indicano un cambiamento all'interno della popolazione nel tempo (e nello spazio), e ciò non permetterebbe di distinguerle in due specie; più che altro, A. anamensis potrebbe rappresentare uno stadio giovanile di A. afarensis.
In parole povere le specie di Australopithecus potrebbero essere 5 (A. afarensis, A. africanus, A. garhi, A. platyops e A. sediba), e potrebbero aumentare se il cosiddetto "Burtele foot" risultasse diverso da quello di Ardipithecus ramidus, oppure se considerassimo anche la morfologia craniodentale (simile a quella di A. afarensis) ed un piede simile a quello di A. ramidus, allora potremmo considerare valida A. deyiremeda. Questo cambierebbe anche il range temporale del genere:
Sulla base dei dati a nostra disposizione, il range temporale del genere Australopithecus si estenderebbe tra i 4,2 e i 2 milioni di anni, con i primi membri di A. afarensis posti "alla base" e gli ultimi individui di A. sediba a "chiudere il cerchio";
Se dovessero essere "convalidate" le 5 specie citate prima, sia l'area geografica che il range temporale sarebbero più ampi.
Filogenesi e biogeografia
In questo paragrafo non avremo una risposta esaustiva dal punto di vista filogenetico, ma ciò che sappiamo è che Australopithecus, a livello temporale, si sviluppa dopo Ardipithecus, mentre Homo e Paranthropus si origineranno da gruppi di australopitecine. I primi fossili associati alle prime australopitecine sono stati rinvenuti presso Asa Issie (Etiopia), attorno ai 4,2 milioni di anni, mentre gli ultimi fossili associati a A. ramidus sono datati a circa 4,3 milioni di anni.
Qui, come vedete, facciamo affidamento esclusivamente sul record fossile, quindi con molta probabilità queste datazioni sono sottostimate: Australopithecus potrebbe essere comparso prima, mentre A. ramidus potrebbe essersi estinto in tempi successivi. Pertanto, non si esclude una sovrapposizione nel tempo di questi due generi.
Anche a livello filogenetico, non dobbiamo pensare all'evoluzione degli ominini nel modo "classico", con l'antenato comune che dà origine a due gruppi separati (speciazione), ma a una sorta di "multi-ramificazione". Ma ci arriveremo tra poco; concentriamoci su alcune morfologie. La differenza tra A. ramidus e Australopithecus riguarda la locomozione, l'habitat e altre morfologie; pertanto, una relazione diretta antenato-discendente è alquanto improbabile. Il tutto viene "confermato" dalla relazione tra Australopithecus e gli altri ominini arcaici, come Orrorin e Sahelanthropus, poiché a livello morfologico le australopitecine possiedono caratteri decisamente diversi.
Per quanto riguarda le discendenze di Homo e Paranthropus da Australopithecus, come accennavamo prima, non dobbiamo pensare a una relazione antenato-discendente proprio perché Australopithecus presenta una diversità morfologica elevata, una distribuzione ampia e una notevole longevità (4,2-2,9 milioni di anni fa circa). A. afarensis potrebbe essere una forma ancestrale di A. africanus, Homo, K. platyops e Paranthropus, con queste "forme" che si sono originate in varie e diverse parti dell'Africa.
A. africanus è probabilmente il risultato di un isolamento geografico di una parte di una popolazione di A. afarensis che si spostò verso sud, attorno ai 3 milioni di anni, con A. africanus che sembra aver dato origine a A. sediba.
Per quanto riguarda Homo, è ancora difficile capire da chi possa essersi originato, anche perché è stato recentemente scoperto che Homo erectus, A. sediba e Paranthropus robustus convissero per un certo periodo. I primi fossili compaiono nel record geologico attorno ai 2,8 milioni di anni, come testimoniato dalla mandibola di Ledi-Geraru, ma la morfologia di questo fossile suggerisce che ci devono essere state modificazioni nette per "passare" da A. afarensis a Homo: il profilo della mandibola più "parabolico", i margini inferiori e alveolari subparalleli rispetto al corpo, il foro mentale che si apre direttamente posteriormente, il ramo ascendente posizionato più posteriormente; i molari più piccoli e più stretti medio-lateralmente, una distinta cuspide sul primo molare (assente in A. afarensis). Insomma, la scarsità fossilifera non aiuta in questo contesto.
Il discorso è leggermente diverso tra A. afarensis e Paranthropus, in quanto, nonostante la diversa morfologia dentaria, esistono delle morfologie in A. afarensis che ricordano quelle specializzate di Paranthropus aethiopicus e P. robustus: una faccia prognatica, una faccia mediana piatta espansa medio-lateralmente, archi zigomatici robusti e grandi molari. Queste morfologie differiscono per la dimensione, rendendo la relazione antenato-discendente più plausibile, oltre a suggerire una diversa biomeccanica della mascella/mandibola e un diverso adattamento alimentare. Ciò ha permesso comunque una distinzione tra i due generi; altrimenti, i parantropi sono conosciuti anche come "australopitecine robuste".
A. garhi possiede denti post-canini molto più grandi e un P3 (premolare superiore) meno asimmetrico, e ciò permetterebbe in primo luogo un discostamento da A. afarensis. Tuttavia, la presenza di caratteri plesiomorfi come una ridotta capacità cranica, una regione sottonasale prognatica e una cresta sagittale, seppur con qualche differenza su certi dettagli, sono condivisi con A. afarensis. Pertanto, anche qui si potrebbe indicare una relazione tra queste due specie come antenato-discendente.
In generale, si indica che A. afarensis fosse una specie longeva e cosmopolita e che abbia dato origine a più taxa in diverse regioni, non attraverso un modello di biforcazione "classico" (un antenato comune che non convive con le due discendenze), ma attraverso una ramificazione "a cespuglio" in cui la specie ancestrale convive con i suoi discendenti.
Breve sintesi delle specie che si sono originate da A. afarensis:
A. africanus e A. platyops (se si dovesse "eliminare" il genere Kenyanthropus) esistevano già più di 3 milioni di anni fa. L'Africa orientale sembra essere stata una sorta di "seconda culla dell'umanità" da cui si sono verificate dispersioni in tempi diversi (e questo varrà anche per Homo sapiens in seguito). Prima dell'inizio della glaciazione dell'emisfero settentrionale, verificatasi attorno ai 3 milioni di anni, il clima caldo-umido del Pliocene ha prodotto ambienti boscosi che hanno creato "gallerie", mettendo in collegamento vari ambienti e rendendo fiumi e laghi interconnessi tra loro in modo continuo. Questo periodo è caratterizzato da una serie di taxa adattati ai boschi e dipendenti dall'acqua, rendendo così isolate le specie adattate ai climi aridi.
Dopo i 3 milioni di anni, in diverse regioni dell'Africa, A. afarensis ha dato origine a Homo, Paranthropus e A. garhi. Il clima non è più caldo e umido, ma diventa più secco e fresco, favorendo l'estensione delle praterie e degli ambienti aperti. Qui la situazione si capovolge: gli organismi adattati agli ambienti forestali e boschivi e dipendenti dall'acqua vengono isolati, mentre gli animali adattati al pascolo, come i bovidi, espandono il proprio areale. Questo cambiamento climatico ha accelerato fenomeni di speciazione nei mammiferi, favorito l'endemismo e la dispersione degli ominini, considerando anche i fenomeni geo-tettonici che hanno "spinto" ulteriormente gli ominini a migrare in varie parti dell'Africa.
Paleoambienti e Paleoecologia
Gli ambienti in cui visse Australopithecus, come Laetoli, Dikika, Hadar, ecc., mostrano una grande variabilità, suggerendo che questo genere si adattò a svariati habitat o, meglio ancora, presentava una serie di caratteristiche funzionali adatte a diversi contesti. A Kanapoi, ad esempio, ci troviamo in una condizione a "mosaico", caratterizzata da forte stagionalità, ricche fonti d'acqua e una moltitudine di specie, sia terrestri che acquatiche. Qui gli ominini avevano l'imbarazzo della scelta, prediligendo sia le risorse terrestri che arboree poiché il sito era anche ricco di predatori, esercitando così una vera e propria pressione selettiva.
Ricerche recenti hanno affermato che Australopithecus fosse adattato agli ambienti stagionali, che cambiavano più volte nel corso di un intero anno, quindi erano in grado sia di vivere in habitat umidi e boschivi che nelle praterie. Un'altra ricerca ha studiato l'abbondanza faunistica delle varie regioni, l'usura dentaria, l'ipsodontia, ed è emerso che ad Allia Bay e Kanapoi erano caratterizzate da un mix di habitat tra cui praterie, boschi e boscaglie.
Australopithecus era un genere flessibile e poco specializzato, il che gli permise di vivere in habitat diversi, dai "campi aperti" a quelli forestali e boschivi. Questa flessibilità probabilmente permise al genere di diversificarsi, sia a livello morfologico che genetico, dando origine a diversi gruppi e/o specie e alla comparsa dei generi Homo e Paranthropus, capaci di sfruttare con successo le diverse risorse trovate in varie condizioni ambientali.
I reperti e i dati provenienti da Mille-Logya e Ledi-Geraru indicano che dopo circa 3 milioni di anni fa le condizioni "aperte" hanno preso il sopravvento su quelle "chiuse" (forestali e boschive), favorendo la comparsa di nuove specie di Australopithecus e le prime associazioni con i generi Homo e Paranthropus, con la probabile estinzione di A. afarensis. Per esempio, il primo individuo associato al genere Homo è stato rinvenuto nel sito di Ledi-Geraru, indicando un momento in cui la variabilità climatica e l'aridità africana hanno innescato vari eventi di speciazione, non solo negli ominini, ma in molti gruppi di mammiferi.
In altri siti come Laetoli, Hadar e Dikika, invece, erano presenti principalmente A. afarensis in ambienti più boschivi e relativamente più umidi. Di conseguenza, Ledi-Geraru e Mille-Logya possono effettivamente essere considerati "la culla del genere Homo e Paranthropus" proprio perché gli habitat più aperti, caratterizzati da erbe e bassi arbusti, sono gli unici che presentano ominini caratterizzati da una grande variabilità morfologica (e non solo!) e sono i primi a restituire fossili appartenenti al nostro genere.
Alimentazione ed uso degli strumenti
Esistono diverse prove che indicano chiaramente che Australopithecus era un genere caratterizzato da una dieta prevalentemente vegetariana, composta da erba, foglie e frutta. Sullo smalto di alcuni denti, avremmo dovuto trovare chiari segni di abrasione, sintomo di una dieta caratterizzata da cibi duri, ma le analisi sulla micro-usura indicano tutt'altro. Infatti, il genere non presenta superfici dentali "bucherellate", come ci si aspetterebbe da un assiduo mangiatore di cibi "duri". Ciò risulta essere "strano" proprio per via della distribuzione geografica del genere e dell'occupazione dei più disparati habitat.
Australopithecus afarensis presenta una vasta gamma di micro-usure dentarie, con una complessità leggermente inferiore a quella di Australopithecus africanus. Tuttavia, intervengono gli isotopi stabili del carbonio che indicano che la dieta era principalmente composta dal consumo di piante C3 (tipiche di ambienti temperati), soprattutto nei primi individui di A. afarensis (o individui di A. anamensis). Dopo circa 3,76 milioni di anni fa, sono state introdotte nella dieta anche piante C4 (tipiche di ambienti caldi con ridotta disponibilità d'acqua), come nel caso di A. africanus, che presenta un'usura dentaria leggermente maggiore a causa di una maggiore introduzione di questi alimenti.
La più recente A. sediba (circa 2 milioni di anni fa), per esempio, consumava più alimenti C3 rispetto a quelli C4, nonostante questi ultimi fossero più disponibili.
Quindi, non solo si nutrivano di cibi relativamente "morbidi", ma ci sono ormai prove dell'utilizzo di strumenti litici per la rimozione della carne e del midollo osseo. Gli strumenti lomekwiani, datati circa 3,3 milioni di anni fa, sono strumenti relativamente meno sofisticati rispetto a quelli Oldowan, ma comunque adempievano al loro scopo e dimostrano che Australopithecus era in grado di produrre strumenti in pietra.
Anche a livello anatomico ci sono prove a sostegno di ciò, poiché A. africanus mostra uno sviluppato osso trabecolare nei metacarpi, simile a quello del genere Homo, indicando una notevole abilità nella manipolazione e nella presa degli oggetti, grazie alla forte opposizione pollice-dito di cui era dotato.
Quindi, utilizzavano strumenti in pietra per integrare nelle loro diete anche risorse animali, almeno occasionalmente, e erano molto flessibili nel consumo, includendo sia prodotti di piante C3 che C4. Aumentavano il consumo di quest'ultima categoria in concomitanza ai cambiamenti ambientali in atto in quel momento, cioè con la formazione di ambienti sempre più "aperti".
Comportamenti e adattamenti locomotori
La morfologia delle pelvi, del basicranio e degli arti inferiori indica che Australopithecus fosse un ominino bipede abituale, anche se l'arto superiore e la spalla pongono qualche problema d'interpretazione. Alcuni ricercatori affermano che sia l'arto superiore sia il tronco mostrino caratteristiche arcaiche tipiche dell'antenato arboricolo, classificando così questo genere come capace sia di arrampicarsi saltuariamente (considerando questo carattere una sorta di "organo vestigiale" privo di funzionalità) sia di camminare in modo regolare.
Altri, invece, indicano che Australopithecus fosse in grado di arrampicarsi regolarmente e che questa caratteristica sia stata preservata, "mantenuta" e selezionata nel corso del tempo, in quanto avrebbe fornito vantaggi in termini di fitness, oppure potrebbe essersi trattata di una caratteristica neutra. Insomma, il dibattito sull'arborealità è ancora aperto, ma possiamo concentrarci sulla relazione tra forma e funzione dell'arto negli ominoidi.
Le caratteristiche arcaiche del genere A. afarensis sembrano essere neutrali poiché non compromettevano la bipedalità, oppure potrebbero essere considerate vestigiali e non svolgere alcuna funzione nell'arrampicata, come ad esempio le dita lunghe degli arti inferiori (come suggerisce il piede della bambina di Dikika, in cui le dita allungate avrebbero potuto svolgere un ruolo nella "spinta" del piede). Si ipotizza che queste caratteristiche arcaiche abbiano avuto funzioni "secondarie", contribuendo ad aumentare le prestazioni fisiche. Ad esempio, la scapola in A. afarensis (molto simile a quella dei gorilla) potrebbe aver permesso l'arrampicata solo per cercare determinati alimenti o per la costruzione di un giaciglio, sia per garantire maggiore sicurezza durante la notte sia per l'allevamento dei cuccioli.
Le proporzioni degli arti in A. africanus e il rapporto di dimensione delle articolazioni degli arti di A. africanus e A. sediba indicano che queste ultime specie mostrano caratteristiche arcaiche più marcate rispetto ad A. afarensis. La presenza di canali semicircolari dell'orecchio interno in A. africanus e A. afarensis, simili a quelli delle scimmie africane, sembra supportare l'importanza dell'arborealità.
In generale, è ampiamente accettato che Australopithecus trascorresse le giornate prevalentemente sul terreno e in posizione bipede, ma la capacità di arrampicarsi sarebbe stata un'ottima strategia di sopravvivenza, sia per sfuggire ai predatori sia per dormire o curare la prole. Questa situazione è analoga a quella degli odierni gorilla e scimpanzé, che camminano sulle nocche quando sono a terra (per la maggior parte del tempo), ma diventano individui arboricoli quando si nutrono.
È interessante considerare anche la condizione di A. garhi, vissuto circa 2,5 milioni di anni fa, poiché presenta un rapporto omero/femore derivato rispetto ad A. afarensis, con il femore più allungato rispetto alle altre specie. Se questo materiale fosse effettivamente classificato come appartenente a questa specie, potrebbe essere considerato una potenziale specie di transizione, poiché le proporzioni degli altri fossili sarebbero intermedie tra A. afarensis e Homo erectus.
I fossili di A. sediba, invece, pur rappresentando la specie più derivata del genere Australopithecus, indicano quanto fosse importante l'arrampicata per questa specie, sebbene insieme alla bipedalità.
Nelle ricerche condotte, così come in quelle precedenti, non si fa mai riferimento alla frequenza con cui Australopithecus utilizzasse le due tipologie di locomozione, né se usasse solo quella bipede. Questo perché è estremamente difficile determinarlo con il materiale fossile a nostra disposizione. Ciò che sappiamo con maggiore certezza riguarda la biomeccanica di A. sediba, che mostra una camminata bipede leggermente diversa da quella delle altre specie e probabilmente utilizzava diverse varianti di locomozione bipede. Tuttavia, le prove fossili limitate non ci consentono di comprendere appieno la diversità locomotoria all'interno del genere Australopithecus.
Dimorfismo sessuale e comportamenti
Alcuni comportamenti etologico-sociali sono stati dedotti dal dimorfismo sessuale e dalle impronte fossili, perché, come ben sapete, il comportamento non si fossilizza. Le dimensioni del corpo e dei canini hanno giocato un ruolo cruciale in questa tipologia di studi, ed è stato dedotto per 𝘼. 𝙖𝙛𝙖𝙧𝙚𝙣𝙨𝙞𝙨 ed 𝘼. 𝙖𝙛𝙧𝙞𝙘𝙖𝙣𝙪𝙨, grazie all'altezza della corona dei canini, che fossero caratterizzati da un sistema di accoppiamento che prevedeva un basso grado di competizione maschio-maschio, ma ciò sembra valere solo per 𝘼. 𝙖𝙛𝙧𝙞𝙘𝙖𝙣𝙪𝙨.
Per 𝘼. 𝙖𝙛𝙖𝙧𝙚𝙣𝙨𝙞𝙨, invece, il discorso è diverso, perché sembrano presentare un livello di dimorfismo elevato: il peso di un maschio era quasi il doppio di quello di una femmina, ed il livello di dimorfismo si collocherebbe tra quello di gorilla e scimpanzé, indicando poliginia ed una competizione maschio-maschio molto intensa. Molti non sono d'accordo, perché le dimensioni corporee potrebbero essere state influenzate da fenomeni ambientali, quali la disponibilità delle risorse e la pressione predatoria, senza contare che le dimensioni della corona del canino potevano svolgere una funzione meccanica nell'occlusione buccale e non una funzione selettivo-sessuale.
Qualche indizio in più proviene dalla bambina di Dikika, un esemplare giovanile che mostra come la formazione dentale in 𝘼. 𝙖𝙛𝙖𝙧𝙚𝙣𝙨𝙞𝙨 fosse rapida, in modo molto simile agli scimpanzé e molto diversa da quella lenta degli ominini associati al genere 𝙃𝙤𝙢𝙤. Ciò è interessante, perché le dimensioni del cervello dell'𝘼𝙪𝙨𝙩𝙧𝙖𝙡𝙤𝙥𝙞𝙩𝙝𝙚𝙘𝙪𝙨 erano relativamente piccole e l'organizzazione sociale simile a quella di gorilla e scimpanzé, indicando che il tasso di formazione del cervello dopo la nascita in 𝘼𝙪𝙨𝙩𝙧𝙖𝙡𝙤𝙥𝙞𝙩𝙝𝙚𝙘𝙪𝙨 non era correlato e influenzato dalle dimensioni del cervello, indicando che erano animali altriciali, cioè individui caratterizzati da una gestazione di breve durata con la prole che, alla nascita, presenta un certo grado di immaturità a livello biologico e fisiologico. Almeno ciò vale fino a circa 3,3 milioni di anni fa.
Fonte: Alemseged, Z. Reappraising the palaeobiology of Australopithecus. Nature 617, 45–54 (2023).
Esplora il mondo delle Scienze Naturali, della Paleontologia, della Genetica e di tante altre fantastiche branche scientifiche.
Articoli
©2025 – All rights reserved | Le immagini presenti su questo sito provengono dal sito stesso (Unsplash), da Pixabay e Pexels (licenza stock free), da Canva (con licenza in abbonamento) e dalle ricerche citate, sempre accompagnate da primo autore e anno.