Evolution never sTOPS!



Sottoclasse Anapsida
Dal punto di vista paleontologico, gli Anapsida costituiscono un gruppo di amnioti primitivi caratterizzati dall’assenza di finestre temporali nel cranio, una condizione considerata ancestrale rispetto a quelle diapside e sinapside. Le prime evidenze fossili di anapsidi risalgono al Paleozoico, con una diversificazione significativa nel Permiano, seguita da una drastica riduzione alla fine proprio del Permiano.
I PRIMI AMNIOTI: INTRODUZIONE
7/27/20255 min leggere
Nel precedente articolo sugli amnioti, abbiamo parlato in linea generale della loro evoluzione, delle loro caratteristiche e, soprattutto, della suddivisione degli amnioti in 4 gruppi (o sottoclassi) in base alla presenza delle finestre post-orbitali o temporali.
Qui ci concentriamo su un gruppo che, tutto sommato, non riserva grosse sorprese: la sottoclasse Anapsida. Quindi parliamo di quel macro-gruppo che comprende tartarughe, testuggini, ecc. A differenza degli altri gruppi, non sono presenti finestre temporali, con le prime forme che comparvero nel Carbonifero superiore. La condizione di "cranio senza finestre" potrebbe indicare che questo gruppo sia la forma ancestrale degli altri amnioti, ma il record fossile indica che è comparso contemporaneamente alle altre sottoclassi e che, nel corso del Permiano, gli anapsidi furono soggetti a una forte radiazione evolutiva/adattativa.
Come in tutte le storie belle e lineari, bisogna fare un appunto sulle finestre temporali. Infatti, secondo uno studio del 2011 (clicca qui), le tartarughe, a livello filogenetico (parentale), sarebbero molto vicine e affini ai diapsidi, cioè quegli organismi che sono caratterizzati da 2 finestre temporali (dinosauri, uccelli, coccodrilli). Grazie a una tecnica che utilizza i microRNA per classificare gli organismi, le tartarughe (in senso lato, per rimanere un po' vaghi) condividerebbero un antenato recente con le lucertole (lepidosauri), e non con i coccodrilli.
Sostanzialmente, questi microRNA svolgono diverse funzioni all'interno dell'organismo (come attivare/disattivare certi geni), e lo studio in questione ha identificato ben 77 nuovi "gruppi" (o famiglie) di questi microRNA che permettono, come detto prima, di capire le diverse relazioni e parentele tra i diversi gruppi. Infatti, "tartarughe" e "lucertole" condividono 4 famiglie di queste molecole, mentre con altri organismi non sono state trovate affinità. La conclusione è che, per convenzione (e per necessità, per non complicare troppo la situazione), non possiamo far altro che considerare le tartarughe separate dai diapsidi, ma nulla toglie che, con molta probabilità, la perdita di finestre temporali (una condizione anapside) potrebbe essere un carattere derivato da un cranio diapside (e non viceversa: è più facile pensare a una perdita di certe morfologie). C’è anche da dire che ci sono state ricerche contrastanti, come questa del 2014, che posiziona comunque le tartarughe tra i diapsidi, ma non si sbilancia troppo verso gli arcosauri o i lepidosauri.
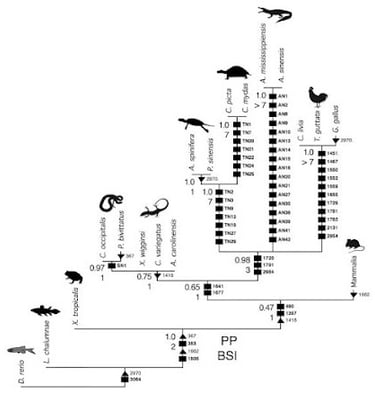
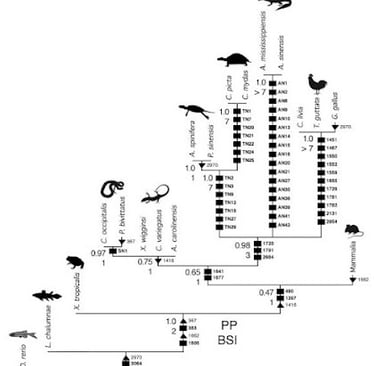
Immagine dove si delinea la parentela tra le tartarughe e gli arcosauri. Fonte: Field J. D. et al., 2014 - appesiaunphylum.com
Un altro lavoro interessante è del 2013, che indica Eunotosaurus africanus, del Permiano (vissuto circa 260 milioni di anni fa), come "parente stretto" delle tartarughe. È uno studio prettamente embriologico incentrato sulla formazione del carapace ed esprime una serie di caratteri rilevanti, tra cui un numero ridotto di vertebre del tronco allungato (nove), nove paia di costole a forma di 'T', una possibile perdita di muscoli intercostali, una riorganizzazione dei muscoli respiratori sul lato ventrale delle costole, ecc. Queste caratteristiche (e altre che potrete trovare nella pubblicazione) sono conformi alla sequenza prevista di "acquisizione del carattere" e forniscono ulteriore supporto al fatto che E. africanus rappresenti una successiva divergenza dal lignaggio delle tartarughe. Quindi, in parole povere, il "modello tartaruga" (inteso come piano corporeo, ecc.) parrebbe essere comparso nel Permiano medio ed è valido per tutti gli 'anapsidi' che possono essere collocati, a livello genetico, esternamente o internamente alla sottoclasse dei diapsidi.
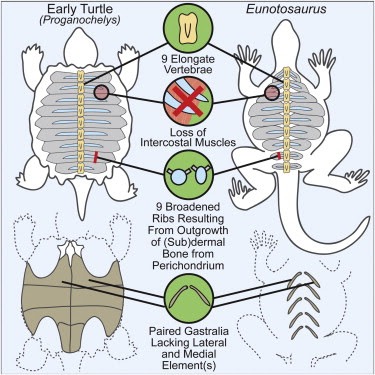
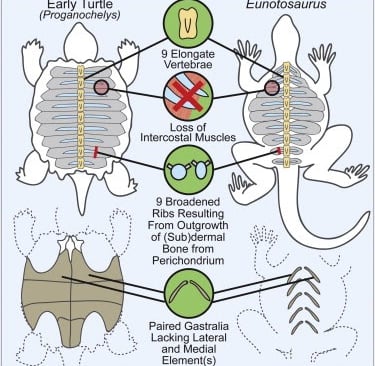
Immagine riassuntiva della ricerca su Eunotosaurus. Fonte: Lyson T. R. et al., 2013 - appesiaunphylum.com
Ma tutto questo è un lavoro per i genetisti e i biologi evolutivi, mentre ora è tempo di virare verso la paleontologia. Come detto prima, gli anàpsidi comparvero nel Carbonifero superiore, ma la maggior parte scomparve alla fine del Permiano. Tra questi gruppi, sopravvissero i Procolofoni (Superfamiglia Procolophonoidea), un gruppo di piccoli animali da cui, secondo alcuni studi, si sarebbero sviluppate le odierne tartarughe. Vediamo qualche caratteristica:
avevano un corpo tozzo, corto, e nel complesso un aspetto simile a quello delle odierne lucertole;
le zampe erano relativamente corte, ma nel complesso sostenevano il corpo;
non superavano i 30 cm di lunghezza;
si nutrivano prevalentemente di insetti o rettili relativamente piccoli.
In parole povere, gli anàpsidi del Permiano erano caratterizzati da un corpo relativamente massiccio, arti sproporzionati e con cinti pelvici abnormi. Quest’ultima struttura, infatti, è lunga come l’omero in tutti i tetrapodi moderni, mentre le proporzioni sono ben diverse in questi primi anàpsidi. Questi gruppi incominciano ad essere diversificati, infatti troviamo per esempio forme erbivore come Scutosaurus karpinskii, una preda tipica dei predatori sinàpsidi.
Non dobbiamo dimenticarci, però, delle fauci di questi organismi. Un anapside primitivo:
possiede tre muscoli principali per chiudere le fauci (adduttore esterno, posteriore e interno, comprendendo anche lo pterigoideo). Nei mammiferi (e nei cinodonti, un antico gruppo di sinàpsidi), per esempio, l’adduttore interno e posteriore sono molto ridotti ed entrano in gioco due nuovi muscoli derivanti da quello esterno;
possiede un muscolo del massetere superficiale e uno temporale in profondità.
Le tartarughe in senso stretto (più o meno) sono comparse e si sono diversificate nel corso del Triassico, probabilmente da qualche "antenato stretto" del gruppo appena citato. Però, già con il genere Proganochelys sp. (per lo studio, clicca qui), incominciamo ad avere un assaggio di prime forme di anapsidi simili alle tartarughe odierne. La pubblicazione è del 1990, ed è sostanzialmente uno studio osteologico che ci fornisce molte informazioni su questo gruppo, in quanto possiede caratteristiche comuni con i cheloni (e altre completamente assenti in quest'ultimo).
Sinapomorfie cheloniane (caratteristiche derivate, tipiche dei cheloni):
Una corazza ossea costituita da un carapace, formato da ossa costali (con costole fuse), ossa neurali con vertebre toraciche fuse. Il piastrone è formato da clavicola, interclavicola e cinque ossa accoppiate/suturate assieme; il piastrone, assieme al carapace, racchiude il cinto pelvico e scapolare.
Il quadrato (un osso del cranio, quello che nei mammiferi si svilupperà come "incudine", uno degli ossicini dell'orecchio) è concavo posteriormente ed esposto lateralmente sulla "guancia".
Sono assenti i postfrontali e postparietali (altre ossa del cranio).
L’osso lacrimale è di piccole dimensioni.
Premascella, mascella e dentale sono privi di denti (edentuli).
Le staffe sono solide (simili a dei bastoncini), senza processi e/o forami.
Oltre a possedere delle sinapomorfie con i cheloni, questo genere possiede anche autapomorfie (caratteristiche presenti solo in questo genere):
Tubercolo ventrale sul basioccipitale;
Una sorta di mazza all’estremità della coda;
Formula delle falangi (manus e pes) di 2-2-2-2-2;
La morfologia degli arti (assieme al contesto deposizionale dove è stato rinvenuto il fossile) indica che questo genere camminasse sul fondo dell’acqua (dolce), ma che non fosse esclusivamente acquatico o terrestre;
...e caratteristiche tipiche degli amnioti primitivi, assenti nelle tartarughe:
Possiede l’osso sovratemporale e quello lacrimale (assieme al dotto lacrimale);
L’articolazione basipterigoidea è mobile;
È assente la parete laterale ossea nell’orecchio medio;
Il processo paraoccipitale dell’opistotico è saldato/attaccato alla scatola cranica solo alla sua estremità distale.


Immagine di Proganochelys quenstedti, © Claire Houck, via Flickr – Licenza CC BY-SA 2.0. - appesiaunphylum.com
Naturalmente, tutto questo è solo un piccolo "racconto" degli anapsidi e, in parte, della loro evoluzione, ma pian piano aggiungerò informazioni che arricchiranno sempre di più la pagina (questo vale per ogni argomento trattato).
Questa volta siamo nel Triassico superiore, e compare l'ordine Testudines. Non entrerò nella diatriba tra testuggini e tartarughe: considero solo l'ordine in questione, "semplificando" con il termine 'tartaruga'.
In linea di massima, parliamo di animali caratterizzati da un cranio privo di finestre temporali, massiccio, privo di denti e semimobile.
Nel Giurassico la diversificazione continua: infatti abbiamo la comparsa dei sottordini Pleurodira e Cryptodira. Le prime sono caratterizzate dalla capacità di ritrarre la testa sotto il carapace piegando lateralmente il collo, mentre le criptodire piegano verticalmente il collo.
Esplora il mondo delle Scienze Naturali, della Paleontologia, della Genetica e di tante altre fantastiche branche scientifiche.
Articoli
©2025 – All rights reserved | Le immagini presenti su questo sito provengono dal sito stesso (Unsplash), da Pixabay e Pexels (licenza stock free), da Canva (con licenza in abbonamento) e dalle ricerche citate, sempre accompagnate da primo autore e anno.